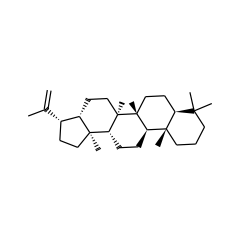
This group of proteins are frequently encoded in the same locus as squalene-hopene cyclase (SHC, IPR006400) and other proteins associated with the biosynthesis of hopanoid natural products. The linkage between SHC and this radical SAM enzyme is strong; one is nearly always observed in the same genome where the other is found. A hopanoid biosynthesis locus was described in Zymomonas mobilis consisting of the genes for HpnA-E and SHC (HpnF). Continuing past SHC are the genes for a phosphorylase enzyme (ZMO0873, i.e. HpnG, IPR017831) and this radical SAM enzyme (ZMO0874) which we name here HpnH. Granted, in Z. mobilis, HpnH is in a convergent orientation with respect to HpnA-G, but one gene beyond HpnH and running in the same convergent direction is IspH (ZM0875, 4-hydroxy-3-methylbut-2-enyl diphosphate reductase), an essential enzyme of IPP biosynthesis and therefore essential for the biosynthesis of hopanoids. One of the well-described hopanoid intermediates is bacteriohopanetetrol. In the conversion from hopene several reactions must occur in the side chain for which a radical mechanism might be reasonable. These include the four (presumably anaerobic) hydroxylations and a methyl shift.
Identification and characterization of Rhodopseudomonas palustris TIE-1 hopanoid biosynthesis mutants
▸ Abstract
Hopanes preserved in both modern and ancient sediments are recognized as the molecular fossils of bacteriohopanepolyols, pentacyclic hopanoid lipids. Based on the phylogenetic distribution of hopanoid production by extant bacteria, hopanes have been used as indicators of specific bacterial groups and/or their metabolisms. However, our ability to interpret them ultimately depends on understanding the physiological roles of hopanoids in modern bacteria. Toward this end, we set out to identify genes required for hopanoid biosynthesis in the anoxygenic phototroph Rhodopseudomonas palustris TIE-1 to enable selective control of hopanoid production. We attempted to delete 17 genes within a putative hopanoid biosynthetic gene cluster to determine their role, if any, in hopanoid biosynthesis. Two genes, hpnH and hpnG, are required to produce both bacteriohopanetetrol and aminobacteriohopanetriol, whereas a third gene, hpnO, is required only for aminobacteriohopanetriol production. None of the genes in this cluster are required to exclusively synthesize bacteriohopanetetrol, indicating that at least one other hopanoid biosynthesis gene is located elsewhere on the chromosome. Physiological studies with the different deletion mutants demonstrated that unmethylated and C(30) hopanoids are sufficient to maintain cytoplasmic but not outer membrane integrity. These results imply that hopanoid modifications, including methylation of the A-ring and the addition of a polar head group, may have biologic functions beyond playing a role in membrane permeability.
Alexander S. Bradley, Ann Pearson, James P. Saenz, Christopher J. Marx
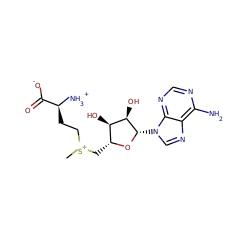
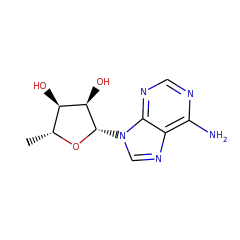
Adenosylhopane: the first intermediate in hopanoid side chain biosynthesis
▸ Abstract
The hopanoid products synthesized by two mutant strains of Methylobacterium together suggest a biosynthetic pathway for the formation of the hopanoid side chain. Mutants deficient in the gene hpnH lack side chains entirely, while those deficient in hpnG accumulate adenosylhopane. These results are in accordance with adenosylhopane as a precursor to extended hopanoids and suggest that adenine is subsequently cleaved, possibly forming phosphoribohopane. We propose that the great diversity of microbial bacteriohopanepolyols and composite hopanoids reflects processes occurring downstream of this intermediate.
Schmerk CL, Welander PV, Hamad MA, Bain KL, Bernards MA, Summons RE, Valvano MA
Elucidation of the Burkholderia cenocepacia hopanoid biosynthesis pathway uncovers functions for conserved proteins in hopanoid-producing bacteria
▸ Abstract
Hopanoids are bacterial surrogates of eukaryotic membrane sterols and among earth's most abundant natural products. Their molecular fossils remain in sediments spanning more than a billion years. However, hopanoid metabolism and function are not fully understood. Burkholderia species are environmental opportunistic pathogens that produce hopanoids and also occupy diverse ecological niches. We investigated hopanoids biosynthesis in Burkholderia cenocepacia by deletion mutagenesis and structural characterization of the hopanoids produced by the mutants. The enzymes encoded by hpnH and hpnG were essential for production of all C35 extended hopanoids, including bacteriohopanetetrol (BHT), BHT glucosamine and BHT cyclitol ether. Deletion of hpnI resulted in BHT production, while ΔhpnJ produced only BHT glucosamine. Thus, HpnI is required for BHT glucosamine production while HpnJ is responsible for its conversion to the cyclitol ether. The ΔhpnH and ΔhpnG mutants could not grow under any stress condition tested, whereas ΔhpnI, ΔhpnJ and ΔhpnK displayed wild-type growth rates when exposed to detergent, but varying levels of sensitivity to low pH and polymyxin B. This study not only elucidates the biosynthetic pathway of hopanoids in B. cenocepacia, but also uncovers a biosynthetic role for the conserved proteins HpnI, HpnJ and HpnK in other hopanoid-producing bacteria.
Environ Microbiol
2015;17(3):735-750
| PubMed ID:
24888970