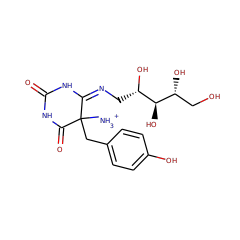
This subunit performs the second half reaction of FO synthase, the rearrangement of 5-amino-5-(4-hydroxybenzyl)-6-((2,3,4,5-tetrahydroxypentyl)amino)dihydropyrimidine-2,4(1H,3H)-dione to form
7,8-didemethyl-8-hydroxy-5-deazariboflavin and ammonia.
Graham DE, Xu H, White RH.
Identification of the 7,8-didemethyl-8-hydroxy-5-deazariboflavin synthase required for coenzyme F(420) biosynthesis
▸ Abstract
The hydride carrier coenzyme F(420) contains the unusual chromophore 7,8-didemethyl-8-hydroxy-5-deazariboflavin (FO). Microbes that generate F(420) produce this FO moiety using a pyrimidine intermediate from riboflavin biosynthesis and the 4-hydroxyphenylpyruvate precursor of tyrosine. The fbiC gene, cloned from Mycobacterium smegmatis, encodes the bifunctional FO synthase. Expression of this protein in Escherichia coli caused the host cells to produce FO during growth, and activated cell-free extracts catalyze FO biosynthesis in vitro. FO synthase in the methanogenic euryarchaeon Methanocaldococcus jannaschii comprises two proteins encoded by cofG (MJ0446) and cofH (MJ1431). Both subunits were required for FO biosynthesis in vivo and in vitro. Cyanobacterial genomes encode homologs of both genes, which are used to produce the coenzyme for FO-dependent DNA photolyases. A molecular phylogeny of the paralogous cofG and cofH genes is consistent with the genes being vertically inherited within the euryarchaeal, cyanobacterial, and actinomycetal lineages. Ancestors of the cyanobacteria and actinomycetes must have acquired the two genes, which subsequently fused in actinomycetes. Both CofG and CofH have putative radical S-adenosylmethionine binding motifs, and pre-incubation with S-adenosylmethionine, Fe(2+), sulfide, and dithionite stimulates FO production. Therefore a radical reaction mechanism is proposed for the biosynthesis of FO.
Critical role of 7,8-didemethyl-8-hydroxy-5-deazariboflavin for photoreactivation in Chlamydomonas reinhardtii
▸ Abstract
DNA photolyases use two noncovalently bound chromophores to catalyze photoreactivation, the blue light-dependent repair of DNA that has been damaged by ultraviolet light. FAD is the catalytic chromophore for all photolyases and is essential for photoreactivation. The identity of the second chromophore is often 7,8-didemethyl-8-hydroxy-5-deazariboflavin (FO). Under standard light conditions, the second chromophore is considered nonessential for photoreactivation because DNA photolyase bound to only FAD is sufficient to catalyze the repair of UV-damaged DNA. phr1 is a photoreactivation-deficient strain of Chlamydomonas. In this work, the PHR1 gene of Chlamydomonas was cloned through molecular mapping and shown to encode a protein similar to known FO synthases. Additional results revealed that the phr1 strain was deficient in an FO-like molecule and that this deficiency, as well as the phr1 photoreactivation deficiency, could be rescued by transformation with DNA constructs containing the PHR1 gene. Furthermore, expression of a PHR1 cDNA in Escherichia coli produced a protein that generated a molecule with characteristics similar to FO. Together, these results indicate that the Chlamydomonas PHR1 gene encodes an FO synthase and that optimal photoreactivation in Chlamydomonas requires FO, a molecule known to serve as a second chromophore for DNA photolyases.
DeCamps L, Philmus B, Benjdia A, White RH, Begley TP, Berteau O
Biosynthesis of F0, precursor of the F420 cofactor, requires a unique two radical-SAM domain enzyme and tyrosine as substrate
▸ Abstract
Cofactors play key roles in metabolic pathways. Among them F420 has proved to be a very attractive target for the selective inhibition of Archaea and Actinobacteria. Its biosynthesis, in a unique manner, involves a key enzyme, F0-synthase. This enzyme is a large monomer in Actinobacteria while it is constituted of two subunits in Archaea and Cyanobacteria. We report here the purification of both types of F0-synthase and their in vitro activities. Our study allows us to establish that F0-synthase, from both types, uses 5-amino-6- ribitylamino-2,4(1H,3H)-pyrimidinedione and tyrosine as substrates but not 4-hydroxylphenylpyruvate as previously suggested. Furthermore, our data support the fact that F0-synthase generates two 5'-deoxyadenosyl radicals for catalysis which is unprecedented in reaction catalyzed by radical SAM enzymes.
J Am Chem Soc
2012;None(None):None-None
| PubMed ID:
23072415
Philmus B, Decamps L, Berteau O, Begley TP
Biosynthetic versatility and coordinated action of 5'-deoxyadenosyl radicals in deazaflavin biosynthesis
▸ Abstract
Coenzyme F420 is a redox cofactor found in methanogens and in various actinobacteria. Despite the major biological importance of this cofactor, the biosynthesis of its deazaflavin core (F0) is still poorly understood. F0-synthase, the enzyme involved, is an unusual multi-domain radical SAM enzyme that uses two separate 5'-deoxyadenosyl radicals to catalyze F0 formation. In this paper, we report a detailed mechanistic study on this complex enzyme that led us to identify (A) the hydrogen atoms abstracted from the substrate by the two radical SAM domains, (B) the second tyrosine-derived product, (C) the reaction product of the CofH catalyzed reaction, (D) the demonstration that this product is a substrate for CofG, and (E) a stereochemical study that is consistent with the formation of a p-hydroxybenzyl radical at the CofH active site. These results enable us to propose a mechanism for F0 synthase and uncover a new catalytic motif in radical SAM enzymology involving the use of two 5'-deoxyadenosyl radicals to mediate the formation of a complex heterocycle.
J Am Chem Soc
2015;137(16):5406-5413
| PubMed ID:
25781338