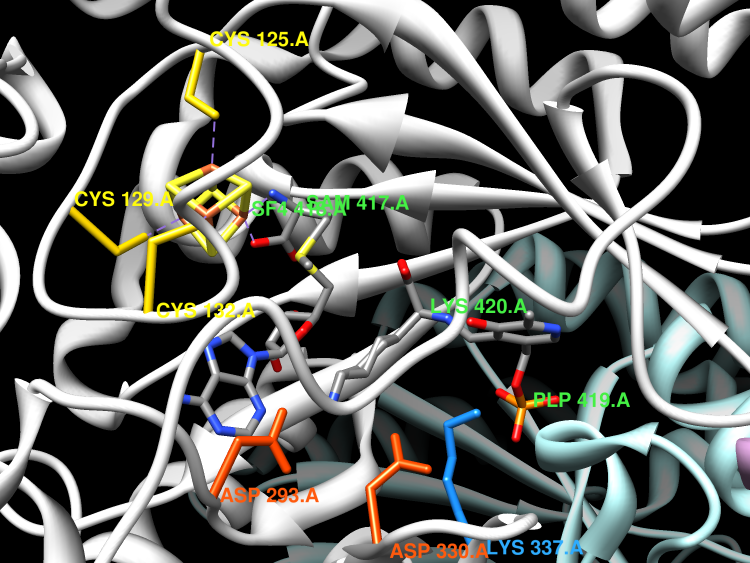
Lysine 2,3-aminomutase (LAM) is a member of the Radical SAM superfamily and is involved in the metabolism of lysine. The superfamily is characterised by the presence of a CxxxCxxC motif which binds a [4Fe-4S] cluster, leaving one of the iron ions unsatisfied. It is often found in anaerobic bacteria that utilise (S)-lysine for growth as a source of nitrogen and/or carbon. Biosynthetically, the (S)-beta-lysine produced is incorporated into antibiotics that contain beta-aminoacyl substituents. Whilst most LAM enzymes produce the (S)-beta-lysine, some produce the R form, such as the Escherichia coli gene yjeK which encodes for a variant of the classical LAM. Whilst zinc is required for enzyme activity,the Zn ion is structural, rather than a cofactor and is required for maintaining the oligomeric state of the biological unit. LAM catalyses the interconversion of L-alpha-lysine and L-beta-lysine, which proceeds via migration of the amino group from C2 to C3 concomitant with cross-migration of the 3-pro-R hydrogen of L-alpha-lysine to the 2-pro-R position of L-beta-lysine. Hydrogen takes place without exchange of solvent protons. The reaction proceeds via several radical intermediates, which are initially generated by the reductive cleavage of the C5'-S bond in SAM.
Lepore BW, Ruzicka FJ, Frey PA, Ringe D
The x-ray crystal structure of lysine-2,3-aminomutase from Clostridium subterminale
▸ Abstract
The x-ray crystal structure of the pyridoxal-5'-phosphate (PLP), S-adenosyl-L-methionine (SAM), and [4Fe-4S]-dependent lysine-2,3-aminomutase (LAM) of Clostridium subterminale has been solved to 2.1-A resolution by single-wavelength anomalous dispersion methods on a L-selenomethionine-substituted complex of LAM with [4Fe-4S]2+, PLP, SAM, and L-alpha-lysine, a very close analog of the active Michaelis complex. The unit cell contains a dimer of hydrogen-bonded, domain-swapped dimers, the subunits of which adopt a fold that contains all three cofactors in a central channel defined by six beta/alpha structural units. Zinc coordination links the domain-swapped dimers. In each subunit, the solvent face of the channel is occluded by an N-terminal helical domain, with the opposite end of the channel packed against the domain-swapped subunit. Hydrogen-bonded ionic contacts hold the external aldimine of PLP and L-alpha-lysine in position for abstraction of the 3-pro-R hydrogen of lysine by C5' of SAM. The structure of the SAM/[4Fe-4S] complex confirms and extends conclusions from spectroscopic studies of LAM and shows selenium in Se-adenosyl-L-selenomethionine poised to ligate the unique iron in the [4Fe-4S] cluster upon electron transfer and radical formation. The chain fold in the central domain is in part analogous to other radical-SAM enzymes.
Proc Natl Acad Sci U S A
2005;102(39):13819-13824
| PubMed ID:
16166264
Ruzicka FJ, Lieder KW, Frey PA
Lysine 2,3-aminomutase from Clostridium subterminale SB4: mass spectral characterization of cyanogen bromide-treated peptides and cloning, sequencing, and expression of the gene kamA in Escherichia coli
▸ Abstract
Lysine 2,3-aminomutase (KAM, EC 5.4.3.2.) catalyzes the interconversion of L-lysine and L-beta-lysine, the first step in lysine degradation in Clostridium subterminale SB4. KAM requires S-adenosylmethionine (SAM), which mediates hydrogen transfer in a mechanism analogous to adenosylcobalamin-dependent reactions. KAM also contains an iron-sulfur cluster and requires pyridoxal 5'-phosphate (PLP) for activity. In the present work, we report the cloning and nucleotide sequencing of the gene kamA for C. subterminale SB4 KAM and conditions for its expression in Escherichia coli. The cyanogen bromide peptides were isolated and characterized by mass spectral analysis and, for selected peptides, amino acid and N-terminal amino acid sequence analysis. PCR was performed with degenerate oligonucleotide primers and C. subterminale SB4 chromosomal DNA to produce a portion of kamA containing 1,029 base pairs of the gene. The complete gene was obtained from a genomic library of C. subterminale SB4 chromosomal DNA by use of DNA probe analysis based on the 1,029-base pair fragment. The full-length gene consisted of 1,251 base pairs specifying a protein of 47,030 Da, in reasonable agreement with 47, 173 Da obtained by electrospray mass spectrometry of the purified enzyme. N- and C-terminal amino acid analysis of KAM and its cyanogen bromide peptides firmly correlated its amino acid sequence with the nucleotide sequence of kamA. A survey of bacterial genome databases identified seven homologs with 31 to 72% sequence identity to KAM, none of which were known enzymes. An E. coli expression system consisting of pET 23a(+) plus kamA yielded unsatisfactory expression and bacterial growth. Codon usage in kamA includes the use of AGA for all 29 arginine residues. AGA is rarely used in E. coli, and arginine clusters at positions 4 and 5, 25 and 27, and 134, 135, and 136 apparently compound the barrier to expression. Coexpression of E. coli argU dramatically enhanced both cell growth and expression of KAM. Purified recombinant KAM is equivalent to that purified from C. subterminale SB4.
Travels with Carbon-Centered Radicals. 5'-Deoxyadenosine and 5'-Deoxyadenosine-5'-yl in Radical Enzymology
▸ Abstract
As a graduate student under Professor R. H. Abeles, I began my journey with 5'-deoxyadenosine, studying the coenzyme B12 (adenosylcobalamin)-dependent dioldehydrase (DDH). I proved that suicide inactivation of dioldehydrase by glycolaldehyde proceeded with irreversible cleavage of adenosylcobalamin to 5'-deoxyadenosine. I further showed that suicide inactivation by [2-(3)H]glycolaldehyde produced 5'-deoxy[(3)H]adenosine, the first demonstration of hydrogen transfer to adenosyl-C5' of adenosylcobalamin. The tritium kinetic isotope effect (T)k was 15, which correlated well with the measurement (D)k = 12 for transformation of [1-(2)H]propane-1,2-diol to [2-(2)H]propionaldehyde by DDH. After establishing my own research program, I returned to the glycolaldehyde inactivation of DDH, showing by EPR that suicide inactivation produced glycolaldehyde-2-yl. In retrospect, suicide inactivation involved scission of adenosylcobalamin to 5'-deoxyadenosine-5'-yl, which abstracted a hydrogen from glycolaldehyde. Captodative-stabilized glycolaldehyde-2-yl could not react further, leading to suicide inactivation. In 1986, my colleagues and I took up the problem of the mechanism by which lysine 2,3-aminomutase (LAM) catalyzes S-adenosylmethionine (SAM) and pyridoxal-5'-phosphate (PLP)-dependent interconversion of l-lysine and l-β-lysine. Because the reaction followed the pattern of adenosylcobalamin-dependent rearrangements, I postulated that SAM might be an evolutionary predecessor to adenosylcobalamin. Testing this hypothesis, we traced hydrogen transfer from lysine through the adenosyl-C5' of SAM to β-lysine. Thus, the 5'-deoxyadenosyl of SAM mediated hydrogen transfer by LAM exactly as in adenosylcobalamin mediated hydrogen transfer in B12-dependent isomerizations. The mechanism postulated that SAM cleaves to form 5'-deoxyadenosine-5'-yl followed by abstraction of C3(H) from PLP-α-lysine aldimine to form PLP-α-lysine-3-yl. PLP-α-lysine-3-yl isomerizes to pyridoxal-β-lysine-2-yl, and a hydrogen abstraction from 5'-deoxyadenosine regenerates 5'-deoxyadenosine-5'-yl and releases β-lysine. Of four radicals in the postulated mechanism, three have been characterized by EPR spectroscopy as kinetically competent intermediates. The analysis of the role of iron allowed researchers to elucidate the mechanism by which SAM is cleaved to 5'-deoxyadenosine-5'-yl. LAM contains one [4Fe-4S] cluster ligated by three cysteine residues. As shown by ENDOR spectroscopy and X-ray crystallography, the fourth ligand to the cluster is SAM, through the methionyl carboxylate and amino groups. Inner sphere electron transfer within the [4Fe-4S](1+)-SAM complex leads to [4Fe-4S](2+)-Met and 5'-deoxyadenosine-5'-yl. The iron-binding motif in LAM, CxxxCxxC, found by other groups in four other SAM-dependent enzymes, is the founding motif for the radical SAM superfamily. These enzymes number in the tens of thousands and are responsible for highly diverse and chemically difficult transformations in the biosphere. Available information supports the hypothesis that this superfamily provides the chemical context from which the much more structurally complex adenosylcobalamin evolved.
Acc Chem Res
2014;47(2):540-549
| PubMed ID:
24308628